Эксперты назвали срок жизни на пенсии будущих пенсионеров :: Экономика :: РБК
Не хуже, чем в 1965-м
Продолжительность жизни 55-летних женщин и 60-летних мужчин в середине 1960-х годов составляла соответственно 24,3 и 15,6 года, указывает в своей статье главный редактор журнала «Демоскоп Weekly», директор Института демографии ВШЭ Анатолий Вишневский. То есть сегодняшние россияне живут на пенсии немногим дольше, чем в 1965 году.
Читайте на РБК Pro
Правительство предусмотрело растянутый до 2028 года для мужчин (до 2034-го для женщин) переход к новым пенсионным возрастам.
Но чтобы избежать «отката назад» и сохранить период жизни на пенсии на уровне 1965 года, ожидаемая продолжительность жизни мужчин, достигших 65 лет, должна увеличиться как минимум на 2,2 года к 2028-му, утверждает Вишневский. Для того же, чтобы не уменьшилось время пребывания на пенсии по сравнению с нынешним, прирост продолжительности жизни 65-летних мужчин в ближайшие десять лет должен быть на уровне 2,7 года.
Ожидаемая продолжительность жизни после 65 лет в России с 1965 по 2015 год выросла лишь на семь месяцев, в то время как во Франции — на 6,8 года, в Италии — на шесть лет, в Испании — на 5,8 года, а в Германии — на 5,7 года, напоминает Вишневский.
Впрочем, авторы доклада ВШЭ не увидели демографических ограничений для повышения пенсионного возраста, указал на презентации замдиректора Института демографии ВШЭ Михаил Денисенко. «Говорить о том, что дожитие низкое, что дожитие препятствует повышению пенсионного возраста, — это совершенно несправедливо. Показатели вписываются в палитру показателей, которые были достигнуты многими странами, когда пенсионный возраст повышался», — сказал он.
«Очевидно, что должны быть разные подходы к мужскому и женскому населению, поскольку уровни дожития разные. Возможно, прирост пенсионного возраста на отдельных этапах должен быть тесно увязан с приростом продолжительности жизни, но это вопросы, которые нуждаются в обсуждении», — добавил эксперт.
О предложении правительства повысить пенсионный возраст с 55 до 63 лет для женщин и с 60 до 65 лет для мужчин премьер-министр Дмитрий Медведев объявил 14 июня. Пенсионная реформа предполагает переходный период. Он начнется с 2019 года и продлится десять лет для мужчин и 16 лет для женщин. Темп увеличения — плюс один год каждый год.
Среди факторов, повлиявших на принятие правительством решения, стало увеличение продолжительности жизни. «В последние годы нам удалось добиться существенного увеличения продолжительности жизни: она достигла почти 73 лет», — сказал Медведев. По данным Росстата, ожидаемая продолжительность жизни при рождении в 2017-м составила 72,3 года. Но при обсуждении изменения возраста выхода на пенсию важен другой показатель — средняя продолжительность предстоящей жизни в том возрасте, в каком назначается пенсия, подчеркивает Вишневский.
Продолжительность жизни: сколько вы проживете?
- Дэвид Шпигельхальтер
- BBC Future
Автор фото, Thinkstock
Подпись к фото,В детстве кажется, что жизнь бесконечна, но с годами убеждаешься в обратном
За столбцами сухих цифр, позволяющих строить прогнозы относительно даты нашей смерти, стоят изменения в обществе, события мирового масштаба и будущие тенденции в сфере старения, подчеркивает корреспондент BBC Future.
Одна из моих статей когда-то была посвящена смерти на рабочем месте, и теперь я, рискуя прослыть фаталистом, решил выяснить, как долго мы проживем.
Говоря об отмеренном нам сроке, мы обычно употребляем термин «средняя продолжительность жизни».
Например, в 89-м псалме (в синодальном переводе Библии – Ред.) говорится, что «дней лет наших — семьдесят лет», хотя до недавних пор дожить и до этого «среднебиблейского» возраста было немалым достижением.
Таблицы Галлея
Некоторым историческим личностям это удалось: первый римской император Октавиан Август преодолел планку в 75 лет, а Микеланджело и вовсе побил все рекорды, успев отметить свое 88-летие.
Еще во времена древних греков зародилось поверье о том, будто возраст, кратный семи, является опасным — особенно «климактерические» 49 и 63 года.
Автор фото, Getty
Подпись к фото,Эдмонд Галлей вычислил период обращения кометы, но не дожил до ее очередного визита в 1758 году
Отчасти для того, чтобы преодолеть это убеждение, в 1689 году один священник из силезского города Бреслау (ныне польский Вроцлав) стал собирать данные о том, в каком возрасте умирали люди.
В итоге накопленные им сведения оказались в Англии, у Эдмунда Галлея, который ненадолго оторвался от процесса открытия новых комет и в 1693 году составил первую в мире серьезную статистическую таблицу продолжительности жизни, использовав оценочные величины ежегодного риска смерти для расчета вероятности дожития до определенного возраста.
Никаких признаков того, будто в возрасте 49 и 63 лет риск смерти повышается, обнаружено не было, так что Галлей развенчал миф об опасности «климактерических лет».
Зафиксированная в таблицах Галлея продолжительность жизни доходила до 84 лет. По его оценкам, вероятность достижения этого возраста составляла 2%, что он и доказал на собственном примере, отпраздновав свой 85-летний юбилей.
При этом он изрядно пережил музыканта Билла Хейли, который также в известном смысле обязан комете своей славой, хотя дотянул только до 55 лет.
 – Ред.
– Ред.Магия (шестидесяти) четырех
На величину продолжительности жизни существенно влияет высокая детская смертность — из-за этого среднее значение получается небольшим, однако те, кто все-таки вступил во взрослую жизнь, могут дожить до весьма преклонных лет.
Рассмотрим еще один пример из музыки, который поможет нам более корректно оценить данные о жизни и смерти.
Автор фото, Getty
Подпись к фото,«Битлз» записали песню «Когда мне будет 64» в 1967 году, Пол Маккартни (крайний слева) сочинил ее в 16 лет
Свой хит «Когда мне будет 64» Пол Маккартни написал в 16 лет — в этом возрасте кажется, будто 64 года — это уже глубокая старость.
Но если сравнить вероятность дожития ребенка до 16 лет с вероятностью достижения им затем возраста 64 лет, мы можем составить себе более правильное представление о том, что изменилось за истекшие десятилетия.
Так, обратившись к столь прекрасным ресурсам, как база данных о смертности человека, мы видим, что в 1841 году (первом, за который имеются данные) 31% детей, родившихся в Англии и Уэльсе, не дожили до 16 лет.
В 1966 году, когда группа «Битлз» записала песню «Когда мне будет 64», в возрасте до 16 лет умерло 2,5% детей, причем у выживших девочек шансы отпраздновать свой 64-й день рождения составляли 85%, а у мальчиков — 74%.
Такая разница объяснялась нездоровым образом жизни многих мужчин.
К 2009 году — последнему, за который зафиксированы данные в этой базе, — до 16 лет не дожили менее 1% детей, а вероятность достижения 64 лет возросла до 92% у женщин и 87% у мужчин.
События мирового масштаба
Однако показатели средней продолжительности жизни имеют свое значение, поскольку за сухими цифрами в статистических таблицах стоят важные события.
Автор фото, Getty
Подпись к фото,Поход Наполеона на Москву сократил среднюю продолжительность жизни на 23 года
Не говоря уже о мировых войнах, на ум приходит поход Наполеона на Москву, унесший жизни 400 000 человек и вызвавший тем самым временное сокращение продолжительности жизни до 23 лет.
Разразившаяся в 1918—1919 годах эпидемия гриппа-«испанки» привела к уменьшению продолжительности жизни француженок на 10 лет.
Результатом распространения СПИДа стало снижение этого показателя в Южной Африке с 63 лет в 1990 году до 54 лет в 2010 году.
В других местах произошли обратные изменения. В 1970 году продолжительность жизни во Вьетнаме составляла 48 лет, а сейчас — 75, тогда как Англии и Уэльсу для совершения подобного скачка потребовалось почти сто лет — с 1894 по 1986 год.
Кроме того, в пределах одной страны мы можем наблюдать последствия неравенства.
Например, в 1901 году в США продолжительность жизни темнокожих мужчин составляла 32 года, а 43% не доживали до 20 лет, тогда как среди белых мужчин эти показатели составляли 48 лет и 24%.
Прошло сто лет, в течение которых активно велась борьба за гражданские права, но разница в продолжительности жизни по-прежнему была заметна, хотя и сократилась с 16 лет до 5.
Старение населения
Нам твердят, что население планеты составляет семь миллиардов человек и стремительно растет. Кроме того, мы можем с разумной долей уверенности строить догадки о продолжительности жизни детей, появившихся на свет за последнее время.
Исходя из того, что в будущем здоровье населения улучшится, можно прогнозировать рост средней продолжительности мужчин, родившихся в Англии и Уэльсе, в среднем до 90 лет, а женщин — до 94.
Автор фото, Thinkstock
Подпись к фото,Хотя население Земли стареет, в некоторых странах людям грозит все меньше старческих недугов
При этом ожидается, что в 2112 году 32% мужчин и 39% женщин отпразднуют свой столетний юбилей.
От таких цифр голова идет кругом. Однако, по вполне понятным причинам, подобные прогнозы содержат в себе некое противоречие.
Есть ли в организме человека встроенный механизм старения, и может ли его действие быть обращено вспять?
Или же существует некий естественный потолок, в который мы и упремся облысевшими от старости головами?
Ясно одно: в будущем дети будут окружены огромным количеством стариков, нуждающихся в уходе, — ведь те, кто сегодня достиг средних лет, отнюдь не собираются умирать.
По оценкам ООН, за период с 2007 по 2050 год доля людей в возрасте старше 60 лет удвоится за счет увеличения продолжительности жизни и снижения фертильности женщин, ведущего к сокращению рождаемости.
К 2050 году в мире будет жить два миллиарда человек в возрасте старше 60 лет и около 400 миллионов старше 80.
Но в каком состоянии будут жить все эти старики? После слов о «семидесяти годах» в 89-м псалме говорится: «а при большей крепости — восемьдесят лет; и самая лучшая пора их — труд и болезнь, ибо проходят быстро, и мы летим».
В общем, счастливой такую старость не назовешь. Неужели все старания продлить свою жизнь приведут лишь к тому, что мы еще дольше просидим в своем уголке, уставившись в телевизор и силясь понять, чего от нас хотят редкие посетители?
На самом деле не всё так плохо. Например, в 2008 году в Великобритании мужчины в возрасте 65 лет могли прожить еще в среднем 17 лет, из которых десять — в «хорошем» или «очень хорошем» состоянии здоровья — а это довольно жесткий критерий.
У женщин в этом возрасте оставалось в запасе более 20 лет, из которых 11 вполне можно было прожить здоровой.
Эти сведения проливают бальзам на мою душу: 64 мне будет через несколько лет, а волос у меня на голове уже почти не осталось.
Но я намерен бороться до конца и еще долго жить у себя дома, а не в богадельне для престарелых любителей статистики.
Названы регионы России с самой высокой ожидаемой продолжительностью жизни
В Ингушетии, по предварительной оценке Росстата, в 2020 году зафиксирована самая высокая ожидаемая продолжительность жизни — 81,2 года. Об этом свидетельствуют данные статистики, с которыми ознакомился RT.
Ожидаемая продолжительность жизни при рождении — это число лет, которое в среднем предстояло бы прожить одному человеку из поколения родившихся в данном году при условии, что на протяжении всей жизни этого поколения уровень смертности в каждом возрасте останется таким, как в год, для которого вычислен показатель.
На втором месте находится Дагестан (76,4 года). Далее в рейтинге идут Москва (76,2), Карачаево-Черкесия (74,8) и Кабардино-Балкария (74,4).
Кроме того, самые низкие показатели зафиксированы на Чукотке — 65,8, в Туве (66,3), Амурской области (67,4), а также в Еврейской автономной области (67,5) и в Забайкальском крае (68,2).
По словам руководителя отдела здоровья и самосохранительного поведения Института демографических исследований ФНИСЦ РАН Аллы Ивановой, в России существует устойчивый региональный вектор изменения продолжительности жизни.
«Продолжительность жизни уменьшается с юга на север и с запада на восток. В основе этого лежат такие фундаментальные причины, которые сформированы ещё в советское время, они продолжают оказывать влияние и в настоящее время. Конечно, главное и основное — развитие социальной инфраструктуры, услуг для населения, уровень жизни населения и социальная структура населения, а также характер занятости. Работа в добывающих отраслях, жизнь в довольно сложных природно-климатических условиях — это всё факторы, которые не способствуют высокой продолжительности жизни», — объяснила она в беседе с RT.
В свою очередь, научный руководитель лаборатории экономики народонаселения и демографии экономического факультета МГУ Валерий Елизаров отметил в разговоре с RT, что можно ещё ожидать небольших изменений показателей в связи с предстоящей переписью населения.
«Что касается этих данных, то ничего удивительного нет, что в верхней части таблицы лучшие показатели имеют Москва и несколько республик Северного Кавказа, а внизу у нас несколько регионов Дальнего Востока, ещё из Сибирского округа Республика Тыва. Все последние годы так, в общем-то, и есть. У нас начиная с 1 октября должна была быть перепись 2020 года, которая уточнила бы эту численность, но она перенесена на апрель… И где-то к концу года мы, может быть, уже получим какие-то предварительные данные в целом по России, чуть позже данные по субъектам Федерации тоже, и вот тогда мы, уже зная уточнённую численность населения и России в целом, и Москвы, и всех субъектов, сможем более корректно посмотреть на данные о смертности 2020 года.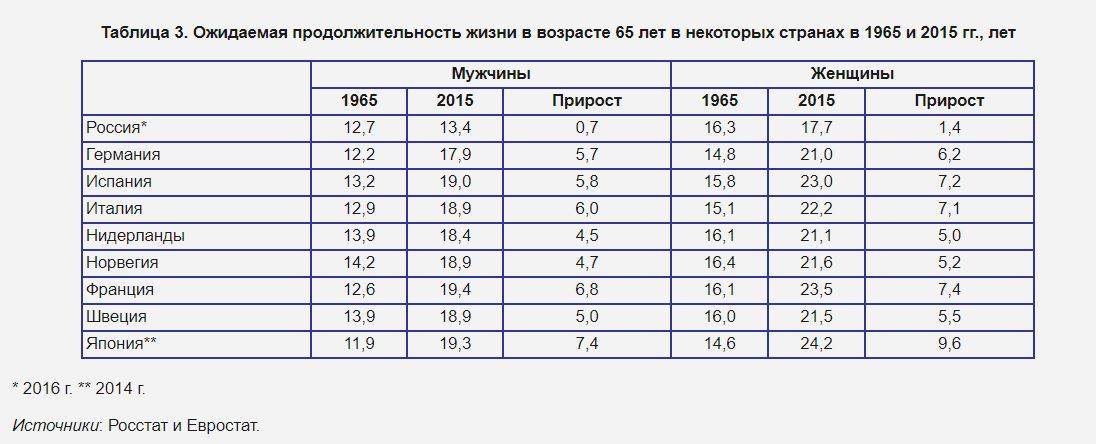 Показатели будут немножко пересчитаны. Я не ожидаю очень большого изменения, но небольшие уточнения будут», — заключил Елизаров.
Показатели будут немножко пересчитаны. Я не ожидаю очень большого изменения, но небольшие уточнения будут», — заключил Елизаров.
В октябре 2020 года российский лидер Владимир Путин на заседании Совета по развитию физической культуры и спорта заявил, что средняя продолжительность жизни в России за десять лет увеличилась более чем на 4,5 года.
Зарядка AirPods и сведения о времени работы от аккумулятора
Узнайте, как заряжать наушники AirPods с помощью зарядного футляра и как продлить время их работы от аккумулятора.
Зарядка наушников AirPods
Чтобы зарядить наушники AirPods, поместите их в футляр. Заряда футляра хватает на несколько полных циклов зарядки наушников AirPods, что позволяет заряжать их в пути. Чтобы наушники AirPods не разряжались, храните их в футляре, когда не используете.
Чтобы наушники AirPods не разряжались, храните их в футляре, когда не используете.
Зарядка наушников AirPods
Можно заряжать зарядный футляр MagSafe или беспроводной зарядный футляр с помощью зарядного устройства стандарта Qi. Расположите футляр на зарядном устройстве так, чтобы его индикатор состояния находился сверху, а крышка была закрыта. Индикатор состояния должен отображать текущий уровень заряда в течение 8 секунд. Если у вас AirPods Pro или AirPods (3-го поколения), можно коснуться футляра, расположенного на зарядном коврике, чтобы увидеть заряжаются ли наушники (индикатор светится оранжевым) или полностью заряжены (индикатор светится зеленым).
Сведения об индикаторе состояния
Зарядный футляр MagSafe: индикатор состояния находится на лицевой стороне футляра.
Беспроводной зарядный футляр: индикатор состояния находится на лицевой стороне футляра.
Зарядный футляр: индикатор состояния в футляре находится между отсеками для наушников AirPods.
Если наушники AirPods находятся в футляре с открытой крышкой, индикатор показывает состояние зарядки. Если наушники AirPods находятся не в футляре, индикатор показывает состояние футляра. Зеленый цвет означает, что аккумулятор полностью заряжен, а оранжевый — что осталось менее одного полного цикла заряда.
Если подключить зарядный футляр MagSafe или беспроводной зарядный футляр к зарядному устройству или поместить его на зарядное устройство стандарта Qi, индикатор состояния будет светиться в течение 8 секунд. Если индикатор мигает белым светом, наушники AirPods готовы к настройке с помощью одного из имеющихся устройств. Если индикатор мигает желтым светом, может потребоваться повторная настройка наушников AirPods.
Если индикатор мигает желтым светом, может потребоваться повторная настройка наушников AirPods.
Чтобы зарядить футляр проводным способом, подсоедините кабель Lightning, входящий в комплект поставки наушников AirPods, к разъему Lightning на футляре. Можно использовать кабель USB-C/Lightning или кабель USB/Lightning. Затем подсоедините другой конец кабеля к зарядному устройству или порту USB. Футляр можно заряжать вне зависимости от наличия в нем наушников AirPods. Быстрее всего это можно сделать, используя зарядное устройство USB для iPhone или iPad или подключив его к компьютеру Mac.
Чтобы просмотреть подробные ведения о состоянии и уровне заряда, см. раздел Проверка состояния заряда.
Проверка уровня заряда аккумулятора в процентах
Уровень заряда аккумулятора наушников AirPods в процентах можно проверить с помощью iPhone, iPad, iPod touch или Mac.
На iPhone, iPad или iPod touch
Откройте крышку футляра, содержащего наушники AirPods, и расположите футляр в непосредственной близости от устройства iPhone, iPad или iPod touch. Подождите несколько секунд, пока на экране не отобразится состояние зарядки наушников AirPods.
Состояние зарядки наушников AirPods можно также проверить с помощью виджета «Элементы питания» на iPhone, iPad или iPod touch. Уровень заряда футляра отображается, только если в футляре находится хотя бы один наушник AirPods.
На компьютере Mac
- Откройте крышку или извлеките наушники AirPods из футляра.

- Щелкните значок Bluetooth () в строке меню.
- Наведите курсор на наушники AirPods в меню.

Сведения о времени работы от аккумулятора
При низком уровне заряда аккумулятора наушников AirPods на экран iPhone или iPad выводится уведомление. Уведомления приходят, когда остается 20, 10 и 5 процентов заряда.
Кроме того, при низком заряде аккумулятора один или оба наушника AirPods воспроизводят звуковой сигнал. При 10 процентах заряда аккумулятора сигнал будет одиночным, а непосредственно перед выключением наушников AirPods — двойным.
Если вы предполагаете проблему с аккумулятором, ознакомьтесь со сведениями по обслуживанию и ремонту наушников AirPods.
Возможности наушников AirPods (3-го поколения)
- Футляр позволяет зарядить наушники несколько раз и слушать музыку до 30 часов,1 а говорить по телефону — до 20 часов. 2.
- AirPods (3-го поколения) могут работать до 6 часов в режиме воспроизведения (до 5 часов с функцией «Пространственное аудио»)3 или до 4 часов в режиме разговора без подзарядки.4.
- Если AirPods (3-го поколения) заряжать в футляре в течение 5 минут, они смогут работать примерно в течение 1 часа в режиме воспроизведения5 или около 1 часа в режиме разговора.6.
 2.
2.Возможности наушников AirPods Pro
- Футляр позволяет зарядить наушники несколько раз и слушать музыку более 24 часов,7, а говорить по телефону — более 18 часов.8
- AirPods Pro могут работать до 4,5 часа в режиме воспроизведения9 или до 3,5 часа в режиме разговора без подзарядки.10.
- Если AirPods Pro заряжать в футляре в течение 5 минут, они смогут работать примерно в течение 1 часа в режиме воспроизведения11 или около 1 часа в режиме разговора. 12.
 12.
12.Возможности наушников AirPods (2-го поколения)
- Футляр позволяет зарядить наушники несколько раз и слушать музыку более 24 часов,13 а говорить по телефону — до 18 часов.14
- Наушники AirPods могут работать до 5 часов в режиме воспроизведения15 или до 3 часов в режиме разговора без подзарядки.16
- Если AirPods заряжать в футляре в течение 15 минут, они смогут работать в режиме воспроизведения до 3 часов17 или до 2 часов в режиме разговора.18
Функция «Оптимизированная зарядка» на AirPods Pro и AirPods (3-го поколения)
Функция «Оптимизированная зарядка» уменьшает износ аккумулятора и продлевает срок его службы за счет сокращения времени, которое наушники AirPods Pro и AirPods (3-го поколения) находятся в полностью заряженном состоянии. Наушники AirPods Pro, AirPods (3-го поколения) и устройства iPhone, iPad и Pod touch на основе анализа вашего ежедневного графика приостанавливают зарядку таким образом, чтобы непосредственно перед использованием наушников был достигнут уровень заряда выше 80 %.
Наушники AirPods Pro, AirPods (3-го поколения) и устройства iPhone, iPad и Pod touch на основе анализа вашего ежедневного графика приостанавливают зарядку таким образом, чтобы непосредственно перед использованием наушников был достигнут уровень заряда выше 80 %.
Для работы функции «Оптимизированная зарядка» AirPods Pro и AirPods (3-го поколения) требуется iPhone, iPod touch или iPad; эта функция включается по умолчанию при настройке устройства либо после обновления iOS или iPadOS до версии 15 или более поздней. Чтобы отключить эту функцию, откройте футляр AirPods и перейдите в меню «Настройки» > Bluetooth на устройстве iPhone, iPad или iPod touch. Нажмите кнопку «Подробнее» рядом с устройством AirPods Pro и AirPods (3-го поколения) в списке устройств. Отключите функцию «Оптимизированная зарядка».
- Тестирование проводилось компанией Apple в сентябре 2021 г. с использованием предсерийных образцов наушников AirPods (3-го поколения) с зарядным футляром MagSafe и ПО, объединенных в пару с iPhone 13 Pro Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %. Тестирование заключалось в полной разрядке аккумулятора AirPods: музыка проигрывалась до остановки воспроизведения на первом наушнике. Разряженные наушники AirPods заряжались на 100 %, а затем музыка снова проигрывалась до остановки воспроизведения на первом наушнике. Цикл повторялся до полной разрядки обоих наушников AirPods и зарядного футляра. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в сентябре 2021 г. с использованием предсерийных образцов наушников AirPods (3-го поколения) с зарядным футляром MagSafe и ПО, объединенных в пару с iPhone 13 Pro Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %. Тестирование заключалось в полной разрядке аккумулятора AirPods: наушники находились в режиме вызова по сотовому телефону до остановки воспроизведения звука на первом наушнике. Разряженные наушники AirPods заряжались на 100 %, а затем снова осуществлялся вызов по сотовому телефону до остановки воспроизведения на первом наушнике. Цикл повторялся до полной разрядки обоих наушников AirPods и зарядного футляра. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в сентябре 2021 г. с использованием предсерийных образцов наушников AirPods (3-го поколения) с зарядным футляром MagSafe и ПО, объединенных в пару с iPhone 13 Pro Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %. Время прослушивания с функциями «Пространственное аудио» и «Отслеживание движения головы» составляло до 5 часов. Тестирование заключалось в полной разрядке аккумулятора AirPods: музыка проигрывалась до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в сентябре 2021 г. с использованием предсерийных образцов наушников AirPods (3-го поколения) с зарядным футляром MagSafe и ПО, объединенных в пару с iPhone 13 Pro Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %. Тестирование заключалось в полной разрядке аккумулятора AirPods: наушники находились в режиме вызова по сотовому телефону до остановки воспроизведения звука на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в сентябре 2021 г. с использованием предсерийных образцов наушников AirPods (3-го поколения) с зарядным футляром MagSafe и ПО, объединенных в пару с iPhone 13 Pro Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %. Тест на 5-минутную зарядку заключался в том, что разряженные наушники AirPods заряжались в течение 5 минут, а затем музыка снова проигрывалась до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в сентябре 2021 г. с использованием предсерийных образцов наушников AirPods (3-го поколения) с зарядным футляром MagSafe и ПО, объединенных в пару с iPhone 13 Pro Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %. Тест на 5-минутную зарядку заключался в том, что разряженные наушники AirPods заряжались в течение 5 минут, а затем снова осуществлялся вызов по сотовому телефону до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в октябре 2019 г. с использованием предсерийных образцов наушников AirPods Pro, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone 11 Pro Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %, и была включена функция активного шумоподавления. Тестирование заключалось в полной разрядке аккумулятора AirPods Pro: музыка проигрывалась до остановки воспроизведения на первом наушнике. Разряженные наушники AirPods Pro заряжались на 100 %, а затем музыка снова проигрывалась до остановки воспроизведения на первом наушнике. Цикл повторялся до полной разрядки обоих наушников AirPods Pro и зарядного футляра. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в октябре 2019 г. с использованием предсерийных образцов наушников AirPods Pro, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone 11 Pro Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %, и была включена функция активного шумоподавления. Тестирование заключалось в полной разрядке аккумулятора AirPods Pro: наушники находились в режиме вызова по сотовому телефону до остановки воспроизведения звука на первом наушнике. Разряженные наушники AirPods Pro заряжались на 100 %, а затем снова осуществлялся вызов по сотовому телефону до остановки воспроизведения на первом наушнике. Цикл повторялся до полной разрядки обоих наушников AirPods Pro и зарядного футляра. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в октябре 2019 г. с использованием предсерийных образцов наушников AirPods Pro, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone 11 Pro Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %, и была включена функция активного шумоподавления. С отключенными функцией активного шумоподавления и Прозрачным режимом время прослушивания составило до 5 часов. Тестирование заключалось в полной разрядке аккумулятора AirPods Pro: музыка проигрывалась до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в октябре 2019 г. с использованием предсерийных образцов наушников AirPods Pro, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone 11 Pro Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %, и была включена функция активного шумоподавления. Тестирование заключалось в полной разрядке аккумулятора AirPods Pro: наушники находились в режиме вызова по сотовому телефону до остановки воспроизведения звука на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в октябре 2019 г. с использованием предсерийных образцов наушников AirPods Pro, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone 11 Pro Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %, и была включена функция активного шумоподавления. Тест на 5-минутную зарядку заключался в том, что разряженные наушники AirPods Pro заряжались в течение 5 минут, а затем музыка снова проигрывалась до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в октябре 2019 г. с использованием предсерийных образцов наушников AirPods Pro, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone 11 Pro Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %, и была включена функция активного шумоподавления. Тест на 5-минутную зарядку заключался в том, что разряженные наушники AirPods Pro заряжались в течение 5 минут, а затем снова осуществлялся вызов по сотовому телефону до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в феврале 2019 г. с использованием предсерийных образцов наушников AirPods (2-го поколения), зарядного футляра, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone XS Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %. Тестирование заключалось в полной разрядке аккумулятора AirPods: музыка проигрывалась до остановки воспроизведения на первом наушнике. Разряженные наушники AirPods заряжались на 100 %, а затем музыка снова проигрывалась до остановки воспроизведения на первом наушнике. Цикл повторялся до полной разрядки обоих наушников AirPods и зарядного футляра. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в феврале 2019 г. с использованием предсерийных образцов наушников AirPods (2-го поколения), зарядного футляра, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone XS Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %. Тестирование заключалось в полной разрядке аккумулятора AirPods: наушники находились в режиме вызова по сотовому телефону до остановки воспроизведения звука на первом наушнике. Разряженные наушники AirPods заряжались на 100 %, а затем снова осуществлялся вызов по сотовому телефону до остановки воспроизведения на первом наушнике. Цикл повторялся до полной разрядки обоих наушников AirPods и зарядного футляра. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в феврале 2019 г. с использованием предсерийных образцов наушников AirPods (2-го поколения), зарядного футляра, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone XS Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %. Тестирование заключалось в полной разрядке аккумулятора AirPods: музыка проигрывалась до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в феврале 2019 г. с использованием предсерийных образцов наушников AirPods (2-го поколения), зарядного футляра, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone XS Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %. Тестирование заключалось в полной разрядке аккумулятора AirPods: наушники находились в режиме вызова по сотовому телефону до остановки воспроизведения звука на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в феврале 2019 г. с использованием предсерийных образцов наушников AirPods (2-го поколения), зарядного футляра, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone XS Max (использовались предварительные версии ПО). Плейлист состоял из 358 уникальных аудиозаписей, приобретенных в магазине iTunes Store (записанных с помощью кодека AAC с битрейтом 256 Кбит/с). Уровень громкости был выставлен на 50 %. Тест на 15-минутную зарядку заключался в том, что разряженные наушники AirPods заряжались в течение 15 минут, а затем музыка снова проигрывалась до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
- Тестирование проводилось компанией Apple в феврале 2019 г. с использованием предсерийных образцов наушников AirPods (2-го поколения), зарядного футляра, футляра с возможностью беспроводной зарядки и ПО, объединенных в пару с iPhone XS Max (использовались предварительные версии ПО). Уровень громкости был выставлен на 50 %. Тест на 15-минутную зарядку заключался в том, что разряженные наушники AirPods заряжались в течение 15 минут, а затем снова осуществлялся вызов по сотовому телефону до остановки воспроизведения на первом наушнике. Время работы от аккумулятора зависит от настроек и характера использования устройства, условий окружающей среды и многих других факторов.
Дата публикации:
Национальный проект «Демография» — Иркутская область. Официальный портал
Реализация национального проекта «Демография» будет способствовать повышению уровня рождаемости, популяризации здорового образа жизни и правильного питания, а также развитию системы долговременного ухода за пожилыми людьми.
Проект направлен на улучшение благосостояния семей за счет мер финансовой поддержки, на увеличение продолжительности здоровой жизни и периода активного долголетия граждан старшего поколения, на увеличение доли регулярно занимающихся спортом граждан.
Цели и целевые показатели на территории России
Обеспечить финансовую поддержку не менее 28 тыс. нуждающихся семей в виде ежемесячной выплаты в связи с рождением первого ребенка.
Провести более 1 тыс. циклов экстракорпорального оплодотворения для семей, страдающих бесплодием.
Увеличить уровень занятости женщин, имеющих детей дошкольного возраста с 64,3% до 68,2%.
Увеличить до 100% доступность дошкольного образования для детей в возрасте от полутора до трех лет.
Не менее 70% лиц старше трудоспособного возраста будут охвачены профилактическими осмотрами и диспансеризацией к концу 2024 года.
Создать гериатрический центр и 2 геронтологических отделения, в которых смогут получить помощь более 2,28 тыс. граждан старше трудоспособного возраста.
Снизить смертность женщин в возрасте от 16 до 54 лет более чем на 10% к уровню 2018 года.
Снизить смертность мужчин в возрасте от 16 до 59 лет более чем на 28% к уровню 2018 года.
Увеличить ожидаемую продолжительность здоровой жизни до 67 лет.
В области в рамках национального проекта реализуется 5 региональных проектов:
Финансовая поддержка семей при рождении детей
Содействие занятости
Спорт — норма жизни
Старшее поколение
Укрепление общественного здоровья
Дата изменения: 16.08.2021 12:11
Доклад о результатах комплексного мониторинга социально-экономического положения пожилых людей в 2019 году
По данным Росстата, в 2019 году в Российской Федерации при незначительном снижении общей численности населения продолжает расти численность лиц старшего возраста.
В 2019 году в Российской Федерации из 146,7 млн человек каждый четвертый находился в возрасте старше трудоспособного (36,6 млн человек), что составляет 25% населения страны.
Прогнозируемая численность населения старше трудоспособного возраста к 2024 году увеличится до 40,8 млн человек (27,6%), а к 2030 году – до 43,7 млн человек. Лица старшего поколения будут составлять более 29% населения страны.
Средняя ожидаемая продолжительность жизни при рождении возросла для мужчин – с 67,75 лет в 2018 году до 68,23 лет в 2019 году, для женщин – с 77,81 лет до 78,17 лет в 2019 году.
Процесс старения населения, выражающийся в возрастании доли граждан старшего поколения в населении страны, вызван снижением суммарного коэффициента рождаемости и ростом продолжительности жизни.
Одной из национальных целей развития Российской Федерации на период до 2024 года, установленных Указом Президента Российской Федерации от 7 мая 2018 г. № 204 «О национальных целях и стратегических задачах развития Российской Федерации на период до 2024 года» (далее — Указ № 204), является повышение ожидаемой продолжительности жизни до 78 лет (к 2030 году — до 80 лет).
Усилия всех структур общества для достижения поставленной цели направлены на оказание системной поддержки гражданам старшего поколения, достижение существенного повышения качества их жизни, посредством решения вопросов, связанных с сохранением здоровья пожилых граждан, социального, экономического и психологического самочувствия и материального благополучия.
Ознакомиться с полным содержанием доклада о результатах комплексного мониторинга социально-экономического положения пожилых людей в 2019 году.doc
Антигистаминные препараты: первое и второе поколения
Аллергия готовься!
Для начала выясним, что способствует развитию аллергической реакции. От чего у нас могут появится внезапный кожный зуд, отек слизистой дыхательных путей, боли в животе и прочие проявления острой аллергии? Виной тому — выброс гистамина, медиатора, который выделяется в ответ на попадание аллергена.
Поэтому в борьбе с проявлениями аллергии важное место занимает прием препаратов, блокирующих рецепторы гистамина, снижающих его влияние на организм. Они называются антигистаминными. Существует два поколения антигистаминных лекарственных препаратов.1, 2 Рассмотрим их подробнее.
Антигистаминные препараты 1 поколения
Каждое их поколение – это своеобразный эволюционный виток в развитии лекарственных средств. Чем они новее, тем благоприятнее их профиль безопасности, риск формирования лекарственной устойчивости ниже, а продолжительность действия выше.2
Первое поколение появилось в начале 20 века, и их эффекты связаны с блокирующим действием на гистаминовые рецепторы. Однако, к сожалению для аллергиков, связь с этими рецепторами очень непрочная и легко обратимая, и именно по этой причине клинический эффект данных препаратов быстро снижается, и требуется частый прием этих лекарств. В среднем, он появляется через 30 минут после приема, а длительность его действия ограничена 4-12 часами. Связано это с тем, что эти препараты быстро метаболизируются и выводятся с мочой.1
К сожалению, у антигистаминных препаратов 1 поколения есть ряд серьезных побочных эффектов, значительно ограничивающих их применение. Дело в том, что действующие вещества проникают через гематоэнцефалический барьер (своего рода физиологический фильтр между кровеносной системой и центральной нервной системой, через который в мозг поступают питательные, биоактивные вещества и часть лекарственных средств), где происходит их связывание с рецепторами головного мозга, развивается блокада центральных серотониновых и м-холинорецепторов. Это может стать причиной развития седативного эффекта разной степени выраженности, от незначительного до сильного.
Есть и другие побочные эффекты — сухость во рту, усиление обструкции (непроходимости) дыхательных путей, повышение вязкости мокроты, тахикардия, нечеткость зрения. Из негативных свойств стоит отметить развитие тахифилаксии, то есть снижения эффекта через определенный промежуток приема, обычно через 15-20 дней.1, 2
Наиболее известные антигистаминные препараты 1 поколения:
-
Димедрол.
-
Мебгидролин.
Может вызывать чувство головокружения, торможение психической активности. Из-за раздражающего влияния на слизистую желудка рекомендуется принимать после еды. -
Хлоропирамин.
Может использоваться при конъюнктивите и аллергии на коже. Для быстрого эффекта назначается в инъекциях.1, 2
Антигистаминные препараты 2 поколения
Препараты 2 поколения отличаются минимальным влиянием на серотониновые и, так называемые, м-холинорецепторы, а также тем, что к ним очень чувствительны гистаминовые рецепторы. Эти препараты действуют продолжительнее — до 48 часов, поэтому их можно принимать реже, что может снизить риск побочных эффектов.1
Они практически не вызывают седативного действия, нет у них и эффекта тахифилаксии, поэтому они могут назначаться длительно (по рекомендации врача). Однако стоит помнить, что препараты 2 поколения могут оказывать нежелательное воздействие на сердце, поэтому при их длительном приеме необходимо контролировать работу сердечной мышцы.1
Антигистаминные препараты второго поколения:
-
Лоратадин
Действует длительно, до 24 часов. Снимает зуд, отеки, нормализует проницаемость капилляров. -
Цетиризин
Применяется для предотвращения и снижения симптомов аллергии. Действует быстро, эффект отмечается примерно через 20 минут и сохраняется больше суток.3 -
Эбастин.
Назначается при проявления аллергии на коже и слизистой глаз. Не проникает в головной мозг, поэтому на нервную систему и психические реакции влияние Эбастина минимально.
Активные метаболиты препаратов 2 поколения
Существуют абсолютно современные разработки, которые практически лишены недостатков лекарств от аллергии предыдущих поколений. Они не влияют на сердце и центральную нервную систему, не подавляют психические реакции, не приводят к привыканию.
К этим препаратам можно отнести фексофенадин и дезлоратадин. Первый показан при сезонном аллергическом рините и при хронической крапивнице, препятствует выбросу гистамина и развитию реакций на аллерген. При этом действие начинается примерно через 1 час после приема дозы, и длится до суток. Второй препарат назначается при аллергическом насморке, крапивнице. Он блокирует выброс биологически-активных факторов аллергии и предупреждает симптомы заболевания. Снимает спазм, уменьшает отеки и зуд, нормализует проницаемость капилляров. Эффект появляется относительно быстро, примерно в течение 30 минут после приема дозы и продолжается до 24 часов.
Сколько длится поколение: измерение разрыва между поколениями
Слово «поколение» часто используется в повседневной беседе. Некоторые люди даже используют генерацию как способ подсчитать течение времени. Однако поколение не так легко измерить по сравнению с десятилетием или столетием.
Что такое поколение и сколько вообще длится поколение? Пытаетесь ли вы прикоснуться к истории своей семьи или хотите узнать, как общество развивалось с течением времени, мы поможем вам.В этой статье мы обсудим все, что вам нужно знать о поколениях.
Что такое поколение?
Термин «поколение» происходит от латинского слова generare, , что означает «порождать». В социальных науках это слово используется для обозначения группы людей, которые родились и жили примерно в одно и то же время.
Это часто используется как способ измерения периодов в истории, потому что большинство людей в одном поколении разделяют множество схожих идей и взглядов, а также были свидетелями одних и тех же событий.Например, большинство американцев, родившихся в 1950-х годах, повзрослели бы достаточно рано, чтобы стать свидетелями и запомнить убийство Кеннеди.
Разные поколения часто имеют разные мировоззрения и карьерные перспективы благодаря уникальным событиям, которые их сформировали. Это один из основных источников напряжения поколений и разногласий в профессиональных или родительско-детских отношениях.
Два типа поколений
В то время как для широкой публики «поколение» имеет только одно значение, в социальных науках есть два способа определения поколений.Эти два типа — семейное поколение и социальное поколение. Чтобы помочь вам понять и отличить их друг от друга, мы разберем их.
Семейное поколение
В физических и биологических науках семейные поколения определяются как группа людей, принадлежащих к одной линии происхождения от предков. Проще говоря, именно так мы обычно определяем поколения. Общепринятая мудрость гласит, что средняя продолжительность семейного поколения составляет четыре поколения в столетие, но эта продолжительность может варьироваться в зависимости от многих факторов.
Одним из определяющих факторов является уровень развития страны. В семьях в развитых странах разрыв между поколениями обычно составляет от 20 до 30 лет. В этих странах люди более не решаются заводить детей в раннем возрасте из-за многих экономических и социальных факторов.
И наоборот, в менее развитых странах разрыв поколений короче, в среднем около 20 лет. Многие люди в развивающихся странах, как правило, выходят замуж в более молодом возрасте, в результате чего следующее поколение рождается раньше.
Социальное поколение
Социальные поколения обычно определяются как «группа людей, которые родились и жили примерно в одно время». Поскольку они пережили похожие события, люди одного поколения обычно имеют много общих черт и мировоззрений.
В западном мире насчитывается восемь самых последних поколений, от Потерянного Поколения, начавшегося в 1883 году, до Поколения Альфа, начавшегося в 2010-х.
Некоторые люди создают новые микропоколения, чтобы объединить поколение, которое имеет частично совпадающие характеристики с предыдущими или последующими поколениями.Например, существует микро-поколение между Millennials и Gen Z , которое называется Zennials.
Сколько длится каждое поколение?
Расчет межпоколенческих интервалов — неточная наука, особенно в отношении семейных поколений. Тем не менее, социальные поколения лучше определены, поскольку они применимы к широкому кругу людей и были зафиксированы в истории.
Если вам интересно, как долго длится социальное поколение, мы разбили восемь западных поколений и сколько лет просуществовало каждое из них.
Утраченное поколение
Иногда известное как «Поколение 1914 года», «Затерянное поколение» составляло основную часть истребителей Первой мировой войны. Большинство экспертов согласились с тем, что это поколение родилось за 17 лет между 1883 и 1900 годами, а затем достигло совершеннолетия во время Первой мировой войны и бурных двадцатых годов.
Наибольшее поколение
Многие представители величайшего поколения или G.I. Поколение — ветераны Великой Отечественной войны. Это поколение состоит из людей, родившихся с 1901 по 1927 год, то есть это поколение просуществовало около 26 лет.
Бесшумное поколение
Это поколение достигло совершеннолетия в послевоенную эпоху. Родившиеся в течение 17 лет между 1928 и 1945 годами, многие из Silent Generation участвовали в Корейской войне, а некоторые также воевали во Вьетнаме.
Поколение бэби-бумеров
Бэби-бумеры получили свое название от послевоенного бэби-бума, который длился 18 лет с 1946 по 1964 год. Бэби-бум сделал это поколение относительно большим, и многие из них либо воевали во Вьетнаме, либо были частью контркультурных движений 60-х.
Поколение X
Поколение X просуществовало 15 лет и состояло из людей, родившихся с 1965 по 1980 год. Это поколение характеризуется как независимое и испытало сильное культурное влияние MTV. Gen X также был свидетелем последних дней коммунизма и увеличения участия матерей в рабочей силе.
Поколение тысячелетия
Названные так, потому что они достигли совершеннолетия на рубеже тысячелетий, миллениалы или поколение Y родились в течение 15 лет между 1981 и 1996 годами.По данным исследовательского центра Pew Research Center, миллениалы обогнали бэби-бумеров, став крупнейшим поколением в Соединенных Штатах.
Поколение Z
Поколение Z, иногда известное как зумеры, состоит из людей, родившихся за 15 лет между 1997 и 2012 годами. Многие из них — дети поколения Y, которых называли «цифровыми аборигенами», поскольку они родились на пике технологического прогресса.
Поколение Alpha
Generation Alpha — первое поколение, полностью рожденное в 21 веке, и одно из последних поколений.Многие эксперты сходятся во мнении, что дети этого нового поколения рождаются в период с начала 2010-х до середины 2020-х годов.
Как долго живут поколения семей?
В то время как историки несут ответственность за определение социальных поколений, область, называемая генетической генеалогией, отвечает за подсчет продолжительности семейных поколений.
Согласно общепринятому мнению, средняя продолжительность жизни одного поколения составляет 25 лет. В примитивных культурах, когда люди спаривались моложе и продолжительность жизни была короче, средняя продолжительность жизни поколения составляла около 20 лет.
Однако исследователи в этой области показали, что более подходящий интервал как для мужских поколений (от отца к сыну), так и для поколений по женской линии (от матери к дочери) больше. Фактический интервал между поколениями мужчин будет на треть больше, а интервал между женскими линиями будет вдвое меньше. Таким образом, средняя мужская линия длится 31-32 года, а женская — 28-29 лет.
Тем не менее, обычная 25-летняя оценка работает просто как оценка.Если между поколениями слишком большой промежуток времени, у исследователей есть повод перепроверить свою информацию, потому что, возможно, были сделаны ошибки по семейным линиям.
Расчет интервала между поколениями вашей семьи
Если вам когда-нибудь было любопытно, как долго просуществовало каждое поколение в вашей собственной семейной линии, вы можете самостоятельно считать интервалы между поколениями ! Все, что вам нужно подготовить, — это даты рождения соответствующих членов семьи — трех поколений достаточно, то есть вас самих, ваших родителей, бабушек и дедушек.Также нужно знать, в каком возрасте у них родились дети.
Расчет интервала одного поколенияЕсли вы пытаетесь вычислить всю длину родословной одного поколения, например, между вами и вашими родителями, вам необходимо усреднить возраст ваших родителей, когда у них были дети. Например, если ваш отец родил вас, когда ему было 27 лет, а вашей матери — 25, средний возраст — 26 лет — это интервал между вами и вашими родителями.
Расчет интервала между поколениями всей семьиЕсли вы хотите узнать, сколько поколений в вашей семье, способ проще.Допустим, вы хотите рассчитать средний интервал между пятью поколениями в вашей семье. Просто вычтите год своего рождения из года рождения самого старшего предка и разделите результат на количество поколений.
Например, ваш самый старший предок родился в 1860 году, а вы — в 2000. Результат будет 140 — разделите это на количество поколений между ними и вами — скажем, 5 — и вы получите 28. Это означает, что средняя продолжительность родословной в вашей семье составляет 28 лет.
Другие расчеты поколенийВы можете применить эти два вычисления разными способами.Например, вы хотите узнать, сколько продлится мужское поколение в вашей семье по сравнению с женским поколением. Вы можете сделать это, рассчитав средние интервалы между поколениями исключительно по мужской линии, а затем сравнив их с интервалом вашей материнской линии.
Заключительные мысли
Нет точного способа превратить поколения в годы. Хотя продолжительность социальных поколений определить легче, для определения продолжительности жизни вашего семейного поколения могут потребоваться дополнительные вычисления.
Изучение прошлых поколений — хороший способ познакомиться с историей своей семьи. Еще один отличный способ помочь в исследовании — просмотреть старые фотографии. Однако они могли быть повреждены после десятилетий хранения — именно здесь на помощь приходит Центр восстановления изображений. Свяжитесь с нами сегодня, чтобы получить фотореставрацию мирового класса по доступной цене!
Важный ориентир для исследований по сохранению — ScienceDaily
Характеристики жизненного цикла — это основные экологические характеристики вида.К ним относятся физические характеристики, такие как масса тела, и физиологические особенности, такие как репродуктивная способность. Экологи исследовали вариации жизненных черт и их роль в определении реакции видов на изменяющиеся условия, такие как изменение климата, а также на антропогенные стрессоры.
Длина поколения — один из наиболее изученных среди таких признаков. Он представляет собой возраст, в котором половину общей репродуктивной продукции достигается человеком. Это фундаментальная информация для популяционной экологии, а также для природоохранной биологии, поскольку она обычно используется в качестве отсчета времени при измерении риска исчезновения видов.Тем не менее, такую важную переменную часто трудно вычислить из-за нехватки подробных репродуктивных данных даже для хорошо изученных групп, таких как млекопитающие.
Недавняя статья, опубликованная в журнале открытого доступа Nature Conservation , представляет собой первую исчерпывающую попытку дополнить базу данных о продолжительности поколений для всех существующих млекопитающих. Эта база данных представляет собой важный ориентир для экологических и связанных с сохранением видов исследований, которые нуждаются в прагматической информации о продолжительности поколения видов.Кроме того, работа обеспечивает методологию расчета продолжительности поколения для видов, когда прямая информация недоступна. Его можно легко распространить на другие таксономические группы, тем самым внося свой вклад в улучшение расширенного набора инструментов, доступного для оценки риска исчезновения, особенно в Красном списке Международного союза охраны природы (МСОП).
База данных о продолжительности поколений была разработана группой глобальной оценки млекопитающих Римского университета Сапиенца (Италия).Лаборатория является партнером Красного списка МСОП, наиболее полной системы, действующей для оценки рисков исчезновения видов (построенной на вкладе тысяч экспертов по всему миру). База данных Красного списка МСОП послужила основой для разработки набора данных о продолжительности поколений млекопитающих.
База данных содержит данные о продолжительности поколения для 5427 видов млекопитающих, а также данные о массе тела взрослых особей, максимальной продолжительности жизни, продолжительности репродуктивной жизни и возрасте при первом воспроизведении. Источник каждой предоставленной информации точно задокументирован.
История Источник:
Материалы предоставлены Pensoft Publishers . Оригинальная история находится под лицензией Creative Commons License. Примечание. Содержимое можно редактировать по стилю и длине.
(PDF) Продолжительность поколения для млекопитающих
Целью этого набора данных было собрать информацию о массе тела для всех млекопитающих на Земле, чтобы мы могли исследовать закономерности массы тела, наблюдаемые в географическом и таксономическом пространстве и во времени эволюции.Нас интересовала наследуемость размера тела по таксономическим группам (насколько сохраняется масса тела в пределах рода, семейства и отряда?), В общей структуре массы тела на разных континентах (остаются ли моменты и другие описательные статистические данные одинаковыми для разных групп). географическое пространство?) и в течение эволюционного времени (насколько быстро модели массы тела повторялись по образцам, наблюдаемым сегодня? Были ли масштабы вымирания плейстоцена специфичными для каждого континента и совпали ли эти события с прибытием человека?).Эти данные также являются частью более крупного проекта, который направлен на интеграцию моделей массы тела в самых разных таксонах (Рабочая группа NCEAS по размеру тела в экологии и палеоэкологии: увязка модели и процесса в пространстве, времени и таксономических масштабах). Мы начали с обновленной версии таксономического списка всех известных современных млекопитающих мира Д. Э. Уилсона и Д. М. Редера (N = 4629 видов), к которому мы добавили оценки статуса, распределения и массы тела, составленные из первичной и вторичной литературы.По возможности мы использовали средние значения массы тела мужчин и женщин, которые, в свою очередь, усреднялись по нескольким местам, чтобы получить значения массы тела нашего вида. Источники представляют собой строчные ссылки в основном наборе данных, а фактические ссылки отображаются в таблице в метаданных. У млекопитающих есть индивидуальные записи для каждого континента, на котором они встречаются. Обратите внимание, что наш набор данных — это больше, чем объединение небольших сборников. Хотя мы в значительной степени полагались на набор данных для рукокрылых К. Э. Джонса (N = 905), справочник CRC по массе тела млекопитающих (N = 688) и набор данных, собранный для Южной Америки П.Марке (N = 505), они составляют менее половины записей в текущей базе данных. Остальные получены из более чем 150 других источников. Кроме того, мы включаем комплексный комплекс видов позднего плейстоцена для Африки, Северной и Южной Америки и Австралии (еще 230 видов). «Поздний плейстоцен» определяется как примерно 11 тыс. Лет назад для Африки, Северной и Южной Америки и как 50 тыс. Лет назад для Австралии, поскольку это время предшествовало антропогенным воздействиям на фауну млекопитающих. Оценки, содержащиеся в этом наборе данных, представляют собой обобщенную ценность вида, усредненную по полу и географическому пространству.Следовательно, эти данные не подходят для постановки вопросов на уровне населения, когда важна интеграция массы тела с конкретными условиями окружающей среды. Включены все существующие отряды млекопитающих, а также несколько архаичных групп (N = 4859 видов). Поскольку некоторые виды обитают более чем на одном континенте (особенно рукокрылые), насчитывается 5731 запись. У нас есть массы тела для следующих видов: Artiodactyla (280 записей), Bibymalagasia (2 записи), Carnivora (393 записи), Cetacea (75 записей), Chiroptera (1071 запись), Dasyuromorphia (67 записей), Dermoptera (3 записи), Didelphimorphia (68 записей), Diprotodontia (127 записей), Hydracoidea (5 записей), Insectivora (234 записи), Lagomorpha (53 записи), Litopterna (2 записи), Macroscelidea (14 записей), Microbiotheria (1 запись), Monotremata ( 7 записей), Notoryctemorphia (1 запись), Notoungulata (5 записей), Paucituberculata (5 записей), Peramelemorphia (24 записи), Perissodactyla (47 записей), Pholidota (8 записей), Primates (276 записей), Proboscidea (14 записей) ), Rodentia (1425 записей), Scandentia (15 записей), Sirenia (6 записей), Tubulidentata (1 запись) и Xenarthra (75 записей).
ПРИМЕЧАНИЕ О ДЛИТЕЛЬНОСТИ ПОКОЛЕНИЯ JSTOR
АбстрактныйВ этой записке рассматривается средняя разница в возрасте между девочкой и ее матерью и средняя разница в возрасте между ней и ее будущими дочерьми. Эти два показателя средней продолжительности поколений различны для стабильной популяции и одинаковы для стационарной популяции. Среднее значение этих двух показателей численно очень близко к тому, которое можно получить по определению Лотки. Однако определение Лотки трудно интерпретировать, а также не существует решения, когда скорость роста равна нулю.Следовательно, среднее значение средней разницы в возрасте между девочкой и ее матерью и между ней и ее дочерьми кажется хорошим определением продолжительности поколения и свободным от недостатков, упомянутых выше. Questa nota esamina la Difference fra l’età media di una ragazza e quella di sua madre e la дифференциация fra l’età media di questa ragazza e quella delle figlie che avrà. Это связано с неправильным интервалом между поколениями и последовательной дифференциацией звука в стабильном и стабильном популярном состоянии.La media di queste due misure è numericamente assai vicina a quella che si otterrebbe application la Definizione di Lotka. Tuttavia, определение Lotka Presenta delle difficoltà di интерпретация inoltre non consente alcuna soluzione quando il tasso d’incremento naturale è uguale a zero. Quindi la media della different fra l’età media di una ragazza e quella di sua madre e fra l’età di quella ragazza e quella delle sue figlie sembra essere una buona definition dell’intervallo medio fra generazioni e non Presenta i Difettititi.Обратите внимание на различие, которое существует между моен-юне-филлом и целуй-де-матерью и отличием в своем сущем-моен-де-сетт-юн-филл и целуй-де-филлес к’ель аура. Ces deux mesures de l’intervalle moyen entre des générations, последовательные sont différentes в соответствии с постоянным населением и стабильным населением. La moyenne de ces deux mesures est numériquement Assez proche de celle qu’on obtiendrait en appliquant la definition de Lotka. Toutefois, определение Lotka présente des затруднения в работе и elle ne согласие en outre aucune solution quand le taux d’accroissement naturel est égal à zéro.C’est pourquoi la moyenne de la différence entre l’âge moyen d’une jeune fille et celui de sa mère et entre l’âge de cette jeune fille et celui de ses filles semble être une bonne définition de l’intervalle moyen qui existe Entre les génerations, sans présenter les défauts cités ci-dessus.
Информация о журналеGENUS — это онлайновый и печатный рецензируемый журнал демографических наук, публикуемый Римским университетом «La Sapienza» при поддержке Департамента статистических наук.GENUS применяет междисциплинарный подход к популяционным исследованиям. Он публикует статьи «чистой» демографии и статьи, направленные на углубление знаний о взаимосвязи между демографией и биологическими, социальными и экономическими явлениями, связанными с эволюцией населения. GENUS всегда пытался продвигать открытый диалог через культурные, идеологические, а иногда и теоретические стены, пытаясь понять сложности, лежащие в основе проблем народонаселения.
Информация об издателеЛа Сапиенца с более чем 700-летней историей и 145 000 студентов является крупнейшим университетом Европы и вторым по величине в мире после Каира.Недавние реформы привели к созданию 11 факультетов. В нем работают более 4500 преподавателей и исследователей, а также около 5000 технических специалистов и административный персонал. La Sapienza предлагает более 370 программ на получение степени, 300 магистров и 117 кандидатов наук. Его услуги включают 155 библиотек, 21 музей, а также справочные и информационные пункты. Его 143 000 студентов включают более 30 000 из-за пределов Рима и более 7 000 не итальянцев, а также около 1000 студентов Erasmus в год. Благодаря 67 отделам и более чем 30 учебным центрам, La Sapienza достигает высокого уровня в различных областях исследований, таких как статистика и демография.Это государственный университет, независимый и бесплатный, стремящийся внести свой вклад в развитие общества знаний посредством исследований, первоклассного обучения и международного сотрудничества.
Китайская онлайн-торговля попугаями: длина поколения и масса тела определяют объем продаж через цену
https://doi.org/10.1016/j.gecco.2020.e01047Получить права и контентОсновные моменты
- •
Масса тела и поколение длина привела объем онлайн-продаж защищенных попугаев над Taobao.com, через влияние на цену.
- •
Онлайн-торговля была наибольшей в отношении попугаев меньшего размера, которые быстрее размножались и, следовательно, дешевле.
- •
При контроле филогенетической зависимости повышение цены на 1% было связано с уменьшением объема продаж на 0,758%.
- •
Более крупные, медленно размножающиеся и более редкие виды могут оказаться под угрозой сохранения, если выловлены в дикой природе.
- •
Более мелкие и более распространенные виды составляют существенную общую проблему благополучия животных.
Реферат
Торговля дикими животными угрожает глобальному биоразнообразию и благополучию животных, где попугаи входят в число наиболее часто торгуемых таксонов, поставляющих экзотических домашних животных и разводящих животных в неволе по всему миру. Используя филогенетический анализ пути, мы исследуем, как биологические факторы взаимодействуют с ценой, чтобы влиять на объемы защищенной онлайн-торговли попугаями в Китае, используя транзакции, зарегистрированные для 46 видов (n = 5862 особи). Наибольшая торговля шла мелкими, быстро размножающимися видами, которые продавались по более низкой цене.Этот ценовой эффект следовал экономическому закону спроса с относительно неэластичным спросом (-0,758), перевешивая такие показатели «качества», как окраска тела и статус сохранности. Мы выделяем две области, вызывающие озабоченность: более крупные, медленно размножающиеся и более редкие виды, даже если они продаются в меньших количествах, могут оказаться под угрозой сохранения, если их собирают в дикой природе. Напротив, чистая численность (более 90% особей были ниже средней длины поколения, массы тела и / или цены) и доступность более мелких и более распространенных видов составляют существенную общую проблему благополучия животных, учитывая, что отлов, ввоз, или разведение в неволе многих видов попугаев в Китае незаконно и, следовательно, не регулируется.Наше исследование подчеркивает важность правильного понимания внутренних отношений между движущими силами торговли дикими животными и растениями для информирования соответствующего руководства.
Ключевые слова
Благополучие животных
Сохранение
Филогенетический анализ пути
Ценовая эластичность спроса
Торговля дикими животными
Рекомендуемые статьиЦитирующие статьи (0)
© 2020 Авторы. Опубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирование статей
Оценка скорости мутации и времени генерации на основе продольных образцов последовательностей ДНК | Молекулярная биология и эволюция
Аннотация
В этой статье мы представляем простой метод оценки скорости мутаций на сайт в год, который также дает оценку длины поколения, когда известна скорость мутаций на сайт в поколении.Оценщик, который использует преимущества полиморфизма ДНК в продольных выборках, является объективным по ряду моделей популяции, включая структуру популяции и изменяющийся размер популяции с течением времени. Мы применяем новый метод к продольному образцу последовательностей ДНК гена env вируса иммунодефицита человека типа 1 (ВИЧ-1) от одного пациента и получаем 1,62 × 10 как частоту мутаций на сайт в год для ВИЧ-1. . Используя независимый набор данных для оценки частоты мутаций на поколение, мы получаем 1.8 дней как продолжительность поколения ВИЧ-1, что хорошо согласуется с недавними оценками, основанными на данных о вирусной нагрузке. Наша оценка времени генерации значительно отличается от недавней оценки Родриго и др. когда используется одна и та же частота мутаций на сайт на поколение. Обсуждаются некоторые факторы, которые могут способствовать различию между разными оценками.
Введение
Скорость мутации на нуклеотид в год является фундаментальной величиной для изучения молекулярной эволюции организма.Скорость мутаций обычно оценивается одним из двух подходов. Первый подход заключается в использовании гомологичных последовательностей ДНК двух видов, время расхождения которых калибруется независимым источником, обычно палеонтологическими данными. Второй подход заключается в непосредственном изучении количества мутаций в одном или нескольких поколениях. Первый подход проще и экономичнее, но применим только тогда, когда доступна надежная оценка времени расхождения. Для сравнения, второй подход в принципе более широко применим, но обычно он требует исследования либо большого количества людей, либо большого сегмента ДНК для получения разумного количества изменений.Также необходимо знать генетическое состояние предка. Для крупных животных использование второго подхода является дорогостоящим из-за длительного времени генерации. Для быстро развивающихся организмов полиморфизмы ДНК в продольных образцах, то есть образцах, взятых в ряд временных точек, обеспечивают еще один способ оценки скорости мутаций.
Время генерации или продолжительность поколения организма — это средний промежуток времени между двумя идентичными и последовательными стадиями жизненного цикла организма.Например, время рождения животных с большим размером тела можно определить как средний промежуток времени, в течение которого взрослый человек производит еще одного взрослого; для вируса, такого как вирус иммунодефицита человека типа 1, время генерации можно определить как средний промежуток времени от высвобождения вириона до того, как он заразит другую клетку и вызовет высвобождение другого вириона. Время генерации не только является частью биологических свойств организма, но также играет важную роль в анализе данных о полиморфизме популяции, поскольку генетические модели популяций обычно разрабатываются с единицами времени, соответствующими определенному количеству поколений, а не дням. или лет.
Жизненный цикл крупных животных можно легко наблюдать, и обычно не составляет проблемы определить время поколения. Например, 20 лет широко используется для одного человеческого поколения. Наблюдать жизненный цикл малых организмов, таких как вирусы, in vivo сложно, поэтому необходимо оценить время генерации. Полиморфизм ДНК в продольном образце дает возможность сделать это, когда доступна независимая оценка частоты мутаций в каждом поколении. Цель этой статьи — представить простой метод оценки скорости мутаций на сайт в год, который также дает оценку времени генерации, если известна скорость мутаций на поколение.Мы применяем метод к продольной выборке от пациента с ВИЧ как для иллюстрации метода, так и для получения оценки частоты мутаций и оценки времени генерации ВИЧ-1. Будут обсуждаться различия между новым методом и методом Родриго и Фельзенштейна (1999).
Теория
Предположим, что образец из n последовательностей взят в момент времени t из популяции гаплоидного организма. Выбор в пользу гаплоидной популяции сделан исключительно для удобства дальнейшего обсуждения, и теория почти идентична для диплоидной популяции.
Пусть d kl будет числом нуклеотидных замен на сайт между последовательностями k и l ( k ≠ l ).\ [\ \ mathit {E} (\ mathit {d_ {kl}}) \ {=} \ {\ theta} _ {\ mathit {t}}, \ (1) \ \]
, где E — ожидание и 𝛉 t — величина, величина которой определяется динамикой населения. Например, это хорошо известно из теории слияния (Kingman 1982 a, 1982 b ; см., E.g., недавний обзор Ли и Фу [1999]), что согласно нейтральной модели Райта-Фишера с постоянным эффективным размером популяции N, значение 𝛉 t равно 2 N μ (например, Tajima 1983), где μ — частота мутаций на сайт в поколении. Как правило, t зависит от времени t , в которое взята проба. Предположим, что вторая проба из m последовательностей была взята через T дней из той же генеральной совокупности.Для последовательностей k и l ( k ≠ l ) из второго образца мы имеем, аналогично уравнению (1), что\ [\ \ mathit {E} (\ mathit {d_ {kl }}) \ {=} \ {\ theta} _ {\ mathit {t} {+} \ mathit {T}}. \ (2) \ \]
Снова по нейтральной модели Райта-Фишера с постоянной эффективной численностью населения размер, 𝛉 т + т = 2 Н мкм. В дополнение к попарному количеству нуклеотидных замен в каждом образце, мы можем исследовать нуклеотидные замены между двумя последовательностями, по одной из каждого образца.\ prime} _ {\ mathit {kl}}) \ {=} \ {\ theta} _ {\ mathit {t}} \ {+} \ {\ mu} \ {\ times} \ (\ mathit {G} \ {\ times} \ \ mathit {T}), \ (3) \ \], где μ — частота мутаций на сайт на поколение, а G — количество поколений в день. Эта взаимосвязь хорошо видна из рисунка 1. Первый член справа от равенства обусловлен тем фактом, что в момент времени t, наследственная последовательность последовательности l является просто другой случайной последовательностью из популяции в момент времени t ; таким образом, ожидаемое количество нуклеотидных замен между этой предковой последовательностью и последовательностью k из образца 1, таким образом, составляет 𝛉 t .Второй член справа от равенства — это ожидаемое количество мутаций на сайт, происходящих в предковой последовательности последовательности k за T дней, при условии, что количество мутаций, происходящих в последовательности в данном количестве GT поколений — переменная Пуассона со средним значением μ GT. Следует отметить, что хотя не каждая последовательность в образце 2 испытала одинаковое количество поколений за последние T дней, уравнение (3) все еще остается в силе, поскольку среднее количество поколений для каждой последовательности составляет G × T .\ prime} _ {\ mathit {kl}}) \ {=} \ {\ theta} _ {\ mathit {t}} \ {+} \ \ mathit {vT.} \ (4) \ \] Пусть Π i будет средним числом нуклеотидных замен на сайт между двумя последовательностями из образца i, , и пусть Π 12 будет средним числом нуклеотидных замен на сайт между двумя последовательностями, одной из образца 1 и одной из образца 2. Тогда, исходя из этих ожиданий, легко увидеть, что несмещенная оценка v равна При дальнейшем предположении, что 𝛉 t одинаково для разных значений t, другая несмещенная оценка v is Хотя оценка v̂ ‘должна иметь меньшую дисперсию, чем v̂, предположение, обеспечивающее ее несмещенность, вероятно, во многих ситуациях сомнительно.Следовательно, v̂ обычно предпочтительнее v̂ ′. Если частота μ мутаций на сайт на поколение известна или была оценена независимо, несмещенная оценка G равна длине L поколения, т. Е. время генерации может быть получено как объединение оценок. Предположим, что были взяты образцы из или различных моментов времени. Для каждой пары выборок можно получить оценку G. Пусть G ij будет оценкой из выборок i, и j. Тогда есть r ( r — 1) / 2 оценки G. Как объединить эти оценки для получения общей оценки G — это не только интересный теоретический вопрос, но и большой практический вопрос. важность. Поскольку каждый G ij является несмещенной оценкой G, для любых весов α ij ≥ 0 и Σ ij α ij = 1, величина также несмещенная. эстимейт г. Следовательно, есть много способов комбинировать попарные оценки. Самый простой метод — взять невзвешенное среднее. То есть, пусть α ij = 2 / [ r ( r — 1)], в результате чего проблема с этой оценкой состоит в том, что члены с большими отклонениями могут доминировать в окончательной оценке. Следовательно, лучший способ объединить оценки — это принять во внимание дисперсию каждой оценки. Пусть T ij будет промежутком времени между отсчетами i и j. Тогда Π ij — Π i отражает в среднем количество мутаций в последовательности в течение периода длиной T ij , что приблизительно равно распределению Пуассона. Следовательно, дисперсия Π ij — Π i должна быть пропорциональна T ij . Фактически, можно показать, что\ [\ Var ({\ Pi} _ {\ mathit {ij}} \ {-} \ {\ Pi} _ {\ mathit {i}}) \ {=} \ \ mathit {c_ {ij}} \ {+} \ \ mathit {vT_ {ij}}, \ (14) \ \]
где c ij — комплексная величина, которая является функцией размеров выборки, T ij , и параметры, которые влияют на динамику популяции, такие как размер популяции, скорость роста и время генерации.Причина, по которой c ij даже зависит от времени генерации, заключается в том, что это зависит от количества наследственных последовательностей, которые имеет образец j -го на момент взятия i -го образца, и это количество зависит от поколения время. Поскольку время генерации — это то, что мы намереваемся оценить, общая оценка G α , которая зависит от времени генерации, нежелательна (хотя дисперсия любого G α зависит от времени генерации).Из-за сложности c ij разумной стратегией при выборе весов является игнорирование как c ij , так и корреляций между оценками. Эта простая стратегия приводит к оптимальным весам α ij = T ij / Σ i < j T ij и соответствующим оценкам v G и где μ — частота мутаций на сайт в поколении.Таким образом, длину L поколения, то есть время генерации, можно оценить как L T = G -1 T .Разница
Дисперсия любой из оценок G α чрезвычайно сложна даже в простой ситуации постоянного размера популяции. Мало того, что формула для отдельного Var ( G ij ) неразрешима, но и различия между значениями G ij коррелированы друг с другом.Поэтому мы предлагаем использовать бутстрап-выборки для оценки дисперсии L T (или G T ).
Пусть n i будет количеством последовательностей в образце i ( i = 1,…, r ). Выборка начальной загрузки будет состоять из r подвыборок, причем i -я подвыборка будет получена путем выбора последовательностей n i с заменой последовательностей n i .Этот стратифицированный бутстрап целесообразен при отсутствии детального знания динамики изучаемой популяции. Начальная оценка дисперсии L T получается с помощью следующих шагов: (1) выполнить выборку начальной загрузки для каждого из образцов r , (2) вычислить значение L T из образцы начальной загрузки, и (3) повторить шаги 1 и 2 много раз; результирующая дисперсия выборки L T является тогда оценкой дисперсии L T .
Следует отметить, что существует два уровня дисперсии, связанных с L T . Один из них связан со стохастичностью эволюции, которая приводит к различиям между репликами одного и того же эволюционного процесса. Другой связан с эффектом выборки. Описанная выше начальная оценка дисперсии учитывает только дисперсию, обусловленную выборкой. Хотя следует проявлять осторожность при интерпретации этого компонента дисперсии, тем не менее, он дает нижнюю границу общей дисперсии.Когда доступно несколько популяций, которые можно рассматривать как реплики одного и того же эволюционного процесса (например, популяции ВИЧ в разных человеческих хозяевах), процедура повторной выборки начальной загрузки, состоящая из выборки как из одной, так и среди реплик, даст оценку общей дисперсии. .
Приложение к ВИЧ
Сообщалось о ряде исследований с участием продольных образцов ДНК-последовательностей вируса иммунодефицита человека типа 1 (ВИЧ-1) внутри хозяина (e.г., Balfe et al. 1990; Simmonds et al. 1991; Wolfs et al. 1991; Холмс и др. 1992; Zhang et al. 1997; Родриго и др. 1999). Мы будем использовать случай, проанализированный Родриго и др. (1999) для иллюстрации нашего метода и с целью сравнения оценок. Этот продольный образец был получен от гомосексуального мужчины европеоидной расы, которому был поставлен диагноз серопозитивного ВИЧ-1 после эпизода асептического менингита в 1985 году (см. Rodrigo et al. [1999] и ссылки в нем). Первый образец был взят в апреле 1989 года, а последующие образцы были взяты через 7, 22, 23 и 34 месяца спустя.Пациент начал лечение зидовудином на 13-м месяце после первой пробы и продолжал до конца исследования.
Область 0,65 т.п.н. гена env
, охватывающая вариабельные области с третьей по пятую, секвенировали для каждого вируса в образце. Мы будем использовать то же выравнивание последовательностей, что и у Rodrigo et al. (1999). Для простоты мы будем рассматривать только нуклеотидные замены. Рассмотрены три метода вычисления числа нуклеотидных замен между двумя последовательностями.Один — это количество нуклеотидных различий, второй — это расстояние с использованием поправки Джукса-Кантора, а третий — это расстояние, скорректированное с использованием двухпараметрической модели Кимуры. Размер выборки и промежутки времени между выборками, а также значения Π для первого и третьего методов приведены в таблице 1.Из таблицы 1 частота мутаций на сайт в день оценивается по формуле v T и составляет 4,71 × 10 -5 без коррекции и 4,45 × 10 -5 с коррекцией с использованием двухпараметрической модели Кимуры.Эти значения соответствуют 1,71 × 10 –2 и 1,62 × 10 –2 на участок в год. Оценка с использованием поправки Джукса-Кантора находится между этими двумя значениями.
Как мы указывали ранее, можно получить оценку времени генерации, если известна частота мутаций на сайт в поколении. Mansky (1996) подсчитал, что общая частота мутаций на сайт на поколение составляла 4 × 10 −5 , включая замену оснований, сдвиг рамки считывания, делецию и вставку.Поскольку в нашем анализе мы рассматриваем только замены оснований, необходимо использовать частоту мутаций только для замен оснований. Используя данные Мански (таблица 4 в Mansky 1996) о 15 заменах оснований в 5272 провирусах челночного вектора с целевым сегментом 114 нуклеотидов, мы получаем степень нуклеотидных замен 15 / (5272 × 114) = 2,5 × 10 −5. на сайт на поколение. Попарные оценки количества G поколений в сутки приведены в таблице 2.
Оценки времени генерации L с использованием исходного расстояния, расстояния с поправкой Джука-Кантора и расстояния с поправкой по двухпараметрической модели Кимуры приведены в таблице 3 вместе с начальными оценками стандартных ошибок.Для сравнения также включены оценки с использованием Rodrigo and Felsenstein (1999) (см. Обсуждение в следующем разделе).
Из таблицы 3 видно, что различия в оценках L среди методов коррекции довольно незначительны, но есть значительные различия между оценками, основанными на G s и G T . Причина, по которой G s почти вдвое больше, чем G T — и, таким образом, G -1 s составляет почти половину G -1 T — это то, что для G s преобладает одна большая оценка G из образцов 3 и 4.Поскольку эти две выборки разделяло всего 28 дней, итоговая оценка G имеет большую дисперсию. Для сравнения, G T дает меньший вес оценке из этих двух выборок, что приводит к меньшему значению и, следовательно, большей оценке времени генерации. Таблица 3 также показывает, что стандартная ошибка начальной загрузки G -1 с немного больше, чем у G -1 T , что дополнительно подтверждает нашу гипотезу, что G −1 T является лучшей оценкой, чем G −1 s .Таким образом, мы заключаем, что время генерации популяции ВИЧ у этого пациента составляет около 1,78 ± 0,25 дня. Для сравнения, комбинированные оценки на основе парных оценок G (таблица 4) методом Родриго и Фельзенштейна (1999) составляют менее половины наших оценок.
Альтернативные методы
Существует несколько существующих методов оценки времени генерации. Здесь мы сосредоточимся на методе, предложенном Родриго и Фельзенштейном (1999), но мы также обсудим другой, совершенно другой подход.Краткое описание метода Родриго и Фельзенштейна (1999) заключается в следующем.
Рассмотрим выборку из m последовательностей из гаплоидной популяции с постоянным эффективным размером популяции N. Если исследовать историю этих m последовательностей, прослеживая назад во времени, можно обнаружить, что существует период, в котором имеется м, наследственных последовательностей, период, в котором имеется м, — 1 наследственных последовательностей и т. д. Временной отрезок t i периода, в котором существует i наследственных последовательностей, имеет экспоненциальное распределение со средним значением, равным 2 N / i ( i — 1) (Kingman 1982 a ).Количество поколений от времени t + T до первого времени выборки t , при котором существует l наследственных последовательностей, составляет G l = t m +… + t l +1 , математическое ожидание которого равно m, — l — это количество событий слияния среди последовательностей m за период времени T. Для пары взятых отсчетов T С интервалом дней можно оценить количество поколений между двумя временами выборки, используя приведенное выше уравнение, если известны как N , так и l .Подобно нашему оценщику, можно преобразовать G l в оценку времени генерации L как T / G l .Эффективный размер популяции N можно оценить исходя из оценки 𝛉 = 2 N μ, где μ — частота мутаций на поколение на сайт. Существует ряд методов оценки 𝛉 (см. Обзор Ли и Фу, 1999 г.). Например, поскольку 2 имеет среднее значение, равное в предположении постоянного эффективного размера популяции, можно оценить N как N̂ = Π 2 / (2μ).Родриго и др. (1999) использовали более сложный метод и получили оценки N , аналогичные оценкам Брауна и Ричмана (1997). Значение m — l было оценено на основе укорененной филогении последовательностей из обеих выборок как количество событий слияния среди последовательностей во второй выборке. Родриго и др. (1999) использовали метод объединения соседей (Saitou and Nei, 1987) для реконструкции филогении и использовали последовательность внешних групп для укоренения дерева. Обратите внимание, что другой древовидный подход для оценки частоты мутаций предложен Рамбо (2000).
Используя подход, описанный выше, Rodrigo et al. (1999) получили оценку времени генерации ВИЧ-1 в 1,2 дня. На первый взгляд их оценка кажется сопоставимой с нашей оценкой в 1,78 дня, но эти две оценки нельзя сравнивать напрямую по двум причинам. Во-первых, Родриго и др. (1999) использовали другой подход, чем наш, для объединения попарных оценок. Во-вторых, мы используем здесь другую частоту мутаций. Хотя рассматривались только нуклеотидные замены, Rodrigo et al.(1999), тем не менее, использовали частоту мутаций 4 × 10 −5 , составленную Mansky (1996), которая включает как инсерции, так и делеции. Когда подсчитываются только нуклеотидные замены, частота мутаций по данным Мански становится равной 2,5 × 10 -5 на сайт на поколение. При такой частоте мутаций попарные оценки G , вычисленные из таблицы 2 Rodrigo et al. (1999) становятся значениями в таблице 4. Сравнение таблицы 4 с таблицей 2 показывает, что оценка Rodrigo et al. (1999) G значительно больше, чем наша, в каждом случае и в среднем более чем в два раза больше, чем наша.Хотя некоторые различия должны быть связаны с расхождениями в обеих оценках, маловероятно, что одни случайные ошибки могут привести к таким систематическим различиям. Некоторые возможные причины этого несоответствия будут обсуждены позже.
Был разработан совершенно иной метод оценки времени генерации с использованием данных продольной вирусной нагрузки внутри хозяина (Coffin 1995; Wei et al. 1995; Perelson et al. 1996). Этот подход основан на том принципе, что когда сильнодействующее лекарство, такое как ритонавир, которое является ингибитором протеазы, вводится пациенту, скорость потери вирионов в плазме может быть смоделирована с помощью набора дифференциальных уравнений с несколькими параметрами. , что можно оценить по данным продольной вирусной нагрузки.Значения этих параметров затем можно использовать для оценки времени генерации. Оценки времени генерации с помощью этого метода варьируются от примерно 4 дней по Wei et al. (1995) до 2,6 дней по Perelson et al. (1996). Последняя группа недавно пересмотрела свою оценку до 1,8 дня (Родриго и др., 1999), что хорошо согласуется с нашей оценкой в 1,78 дня.
Обсуждение
Очень часто статистический метод анализа выборки населения разрабатывается в рамках конкретной модели, такой как постоянный эффективный размер совокупности, принятый Rodrigo et al.(1999). Когда рассматриваемая популяция развивается способом, который значительно отличается от модели, статистический анализ и полученные в результате выводы могут вводить в заблуждение. Поэтому важно понимать, как оценщик ведет себя в различных ситуациях. Предложенная в этой статье оценка частоты мутаций отличается тем, что она беспристрастна в различных ситуациях, что заслуживает дальнейшего обсуждения.
В случае роста населения или, в общем, изменения эффективного размера популяции, легко увидеть, что v ̄ является несмещенной оценкой v , потому что уравнения (5) и (7) выполняются независимо от значения эффективная численность населения.Это свойство нашей оценки особенно важно для его применения к быстро меняющимся вирусным популяциям, таким как ВИЧ-1, потому что популяция внутри хозяина может резко измениться в размере за короткий период времени.
Оценщик v̄ также несмещен при наличии структуры популяции, потому что независимо от структуры популяции каждая последовательность во второй выборке испытывает одинаковое количество времени с момента, когда была взята первая выборка. Пока используется последовательная стратегия выборки из разных выборок, уравнения (5) и (7) остаются в силе независимо от структуры популяции.
Также очевидно, что рекомбинация не вносит смещения в нашу оценку v ̄ , потому что уравнения (5) и (7) справедливы при наличии рекомбинаций. Конечно, это не означает, что непостоянный эффективный размер популяции, структура популяции и рекомбинация не влияют на нашу оценку, потому что они действительно влияют на дисперсию оценки.
Естественный отбор — важный фактор, который следует учитывать при анализе образцов вирусных популяций, таких как ВИЧ-1.Когда исследуемый участок ДНК не участвует напрямую в естественном отборе, наша оценка должна оставаться почти беспристрастной. Это включает ситуацию, в которой исследуемый регион тесно связан с локусом, который находится под сильным естественным отбором. Например, если естественный отбор привел к фиксации благоприятной мутации до начала выборки, то его эффект очень похож на эффект растущей популяции и, следовательно, не приведет к смещению в нашей оценке. Когда многие мутации в образцах не являются селективно нейтральными, накопление нуклеотидных изменений в последовательности за определенный период может отклоняться от распределения Пуассона, и скорость замен может быть увеличена или уменьшена в зависимости от типа естественного отбора.В случае вредных мутаций частота мутаций за год, оцененная из уравнения (13), вероятно, будет меньше, чем экстраполированная частота мутаций на сайт за поколение и время поколения. Это приведет к завышению оценки времени генерации. С другой стороны, если задействован положительный отбор, скорость замены на сайт в год будет повышена, что приведет к недооценке времени генерации. Один из способов минимизировать эффект естественного отбора — проводить анализ только синонимичных замен.Поскольку имеется все больше и больше данных, такой анализ должен быть очень информативным. Поскольку наш анализ в этой статье проводится в основном с целью иллюстрации, а также из-за относительно небольшого размера выборок, мы не проводим более подробный анализ.
Поскольку количество мутаций, существенно повышающих выживаемость вируса, должно быть небольшим по сравнению с общим количеством мутаций, смещение в нашей оценке против из-за положительного отбора вряд ли будет существенным. Тем не менее, поскольку положительный отбор, вероятно, действует на ген env ВИЧ-1 (e.g., Bonhoeffer, Holmes и Nowak, 1995; Ямагути и Годжобори 1997; Zhang et al. 1997), наша оценка времени генерации может немного измениться. Другой потенциальный источник ошибок в оценке времени генерации — это частота мутаций на сайт в поколении. Например, если скорость мутации недооценена, это приведет к недооценке времени генерации. С этими оговорками обнадеживает то, что наша оценка времени генерации хорошо согласуется с недавней оценкой 1.8 дней с момента получения данных о вирусной нагрузке (см. Rodrigo et al. 1999). Будет интересно посмотреть, сохранится ли соглашение с растущими данными.
Что заставляет Родриго и др. (1999) оценивать G , что они постоянно превышают наши? Хотя отклонения в обеих оценках могут привести к колебаниям, расхождение, вероятно, связано с некоторыми фундаментальными различиями между двумя оценками. Одно сходство между двумя оценками времени генерации состоит в том, что обе основаны на оценках количества поколений в день.Однако наш метод является более прямым и беспристрастным для оценки количества поколений в день, в то время как метод Родриго и Фельзенштейна (1999) является косвенным, опираясь на оценки как эффективного размера популяции, так и количества сливающихся событий среди последовательностей в выборке. 2 в периоде, разделяющем две выборки. Подсчет сливающихся событий непосредственно из предполагаемой филогении, вероятно, приведет к завышению этого числа, даже если филогения полностью реконструирована. Простой анализ ниже покажет, почему это так.
Рассмотрим две случайные выборки размером n, и m, соответственно, взятые одновременно. Их слияние будет таким же, как для одного образца из n + m последовательностей. Событие слияния будет засчитываться как слияние последовательностей из образца 2 или просто как слияние внутри образца 2, если и только если ни одна из двух сливающихся последовательностей не имеет потомка в образце 1. Пусть p ( n, m ) — вероятность отсутствия слияния в образце 2.Каждый раз, когда происходит слияние, вероятность выбора каждой пары последовательностей одинакова. Существует n ( n -1) / 2 способов объединения двух последовательностей из образца 1, и есть нм способов объединения одной последовательности из образца 1 и одной последовательности из образца 2. Таким образом, мы имеем следующее Повторяющееся уравнение: Когда есть только одна последовательность во втором образце, нет шансов на коалесценцию в образце 2. Следовательно, начальное условие для решения вышеуказанного повторяющегося уравнения —Вероятность того, что есть по крайней мере одна коалесценция в образце 2 составляет 1- p ( n, m ), а его числовые значения для ряда комбинаций размеров выборки приведены в таблице 5.Из таблицы 5 видно, что для этих размеров выборки в наших продольных выборках эта вероятность довольно высока. Например, когда n = 15 и m = 13, вероятность равна 0,94. Этот анализ предполагает, что даже когда нет времени ( T = 0), разделяющего две выборки, весьма вероятно, что будут наблюдаться слияния во второй выборке, так что подсчет этих событий как оценка м — л , а затем использование этой оценки в качестве основы для оценки G приведет к завышению оценки на G. Это, вероятно, одна из причин, почему оценки Rodrigo et al. (1999) в размере G (таблица 4) неизменно превышают наши оценки (таблица 2). Такое несоответствие также будет наблюдаться, если эффективный размер популяции N недооценен в Rodrigo et al. (1999).
Таблица 5 также показывает, что, когда n велико, 1- p ( n, m ) становится маленьким, что говорит о том, что можно уменьшить смещение в методе Родриго и Фельзенштейна (1999), используя много больший размер выборки для первой выборки.Однако, когда берется более двух образцов, таких как образцы, проанализированные в этой статье, нелегко убедить экспериментатора, скажем, уменьшать вдвое предыдущий размер образца всякий раз, когда берется новый образец, потому что образцы обычно не собираются полностью. для единственной цели. В общем, увеличение всех размеров выборки повысит точность окончательной оценки, что также верно для метода Родриго и Фельзенштейна (1999), потому что время слияния меньше, когда имеется много последовательностей; таким образом, ошибка из-за неправильной оценки -1 не такая серьезная, как для небольшой выборки.
Стоит подчеркнуть, что продольные образцы, необходимые для оценки скорости мутаций и времени генерации, не обязательно должны происходить из одного хозяина. Оценщик применим к выборкам, в которых каждая последовательность исходит от другого хоста. Эта функция полезна для изучения эволюции патогена, который не остается в одном хозяине в течение длительного периода. С другой стороны, если доступны продольные выборки из нескольких популяций, которые можно рассматривать как копии одного и того же эволюционного процесса, точность оценки времени генерации может быть существенно улучшена, поскольку выборки из разных хозяев должны быть более или менее независимыми.Кроме того, общая дисперсия оценки может быть получена путем бутстрэпинга как внутри населения, так и между популяциями. Накапливаются продольные образцы из популяций ВИЧ внутри хозяина, и будет интересно посмотреть, насколько изменчивым может быть время генерации у разных хозяев.
До сих пор мы рассмотрели два способа использования попарного числа нуклеотидных замен. Другое возможное использование уравнений (5) и (7) состоит в оценке отрезка времени T , разделяющего две выборки, когда известны как время генерации, так и частота мутаций на поколение.Оценка T — Потенциальное использование такой оценки — датировать наследственную популяцию, из которой был получен образец древней ДНК. Это, конечно, требует, чтобы современный образец был взят из популяции, которая имела ту же предковую популяцию, из которой был получен древний образец ДНК.Таблица 1 Средние числа ({Π} ij ) (× 10 2 ) нуклеотидных замен
Таблица 1 Средние числа ({Π} ij ) (× 10 2 ) нуклеотидных замен
Таблица 2 Парные оценки G с использованием двухпараметрической модели Кимуры с {μ} = 2.5 × 10 −5
Таблица 2 Парные оценки G с использованием двухпараметрической модели Кимуры с {μ} = 2,5 × 10 −5
Таблица 3 Оценка времени генерации L
Таблица 3 Оценка времени генерации L
Таблица 4 Попарные оценки G Родриго и др. (1999) с {μ} = 2,5 × 10 −5
Таблица 4 Попарные оценки G , выполненные Rodrigo et al.(1999) с {μ} = 2,5 × 10 −5
Таблица 5 Вероятность (1 — p ( n, m )) наличия хотя бы одной слияния среди последовательностей во второй выборке, когда T = 0
Таблица 5 Вероятность (1 — p ( n, m )) наличия хотя бы одного слияния среди последовательностей во втором образце, когда T = 0
Рис. 1. — Схематическая взаимосвязь между двумя последовательностями, взятыми с интервалом T дней
Рис.1. — Схематическая взаимосвязь между двумя последовательностями, взятыми с интервалом T дней
Я благодарю доктора Стэнли Сойера и рецензентов за их комментарии, а доктора Аллена Г. Родриго за любезное предоставление мне своего выравнивания последовательностей. Эта работа была поддержана грантами NIH R29 GM50428 и R01 HG01708, а также стипендией Японского общества содействия науке. Особая благодарность доктору Наруе Сайто за то, что он принял меня в Мисиме.
цитированная литература
Балф, П., П. Симмондс, К. А. Лудлам, Дж. О. Бишоп и А. Дж. Браун.
1990
. Одновременная эволюция вируса иммунодефицита человека типа 1 у пациентов, инфицированных из одного источника: скорость изменения последовательности и низкая частота инактивирующих мутаций.J. Virol.
64
:6221
–6233.Бонхёффер, С., Э. К. Холмс и М. А. Новак.
1995
. Причины разнообразия ВИЧ. Nature 376 : 125Браун, А. Дж. Л. и Д. Д. Ричман.
1997
.ВИЧ-1: игра на эволюции лекарственной устойчивости? Nat. Med. 3 : 268Гроб, Дж. М.
1995
. Динамика популяции ВИЧ in vivo: значение для генетической изменчивости, патогенеза и терапии. Science 267 : 483–489Холмс, Э. К., Л. К. Чжан, П. Симмондс, К. А. Лудлам и А. Дж. Браун.
1992
. Эволюция конвергентной и дивергентной последовательности в гликопротеине поверхностной оболочки вируса иммунодефицита человека типа 1 у одного инфицированного пациента.Proc. Natl. Акад. Sci. USA 89 : 4835–4839Kingman, J. F. C. 1982 a. О генеалогии больших популяций. J. Appl. Вероятно. 19A : 27–43
———. 1982 г. Коалесцентный. Стохаст. Приложения процессов 13 : 235–248
Li, W. H., and Y. X. Fu.
1999
. Теория объединения и ее приложения в популяционной генетике. Стр. 45–79 в Э. Халлоран и С. Гейссер, ред. Статистика в генетике.Спрингер, Нью-Йорк.Манский, Л. М.
1996
. Скорость прямой мутации вируса иммунодефицита человека типа 1 в линии Т-лимфоидных клеток. AIDS Res. Гм. Ретровирусы 12 : 307–314Перельсон, А.С., А.У. Нойман, М. Марковиц, Дж. М. Леонард и Д. Д. Хо.
1996
. Динамика ВИЧ-1 in vivo: скорость клиренса вирионов, продолжительность жизни инфицированных клеток и время генерации вируса. Наука 271 : 1582–1586Rambaut, A.
2000
. Оценка скорости молекулярной эволюции: включение не одновременных последовательностей в филогенетику максимального правдоподобия.Биоинформатика 16 : 395–399Родриго А. Г. и Дж. Фельзенштейн.
1999
. Объединяющие подходы к генетике популяций ВИЧ. В К. А. Крэндалл, изд. Эволюция ВИЧ. Издательство Университета Джона Хопкинса, Балтимор, Мэриленд,Родриго А.Г., Э.Г. Шпаер, Э.Л. Делварт, А.К. Н. Иверсен, М. В. Галло, Дж. Брояч, М. С. Хирш, Б. Д. Уокер и Дж. И. Маллинс.
1999
. Сводные оценки времени генерации ВИЧ-1 in vivo. Proc. Natl.Акад. Sci. USA 96 : 2187–2191Saitou, N., and M. Nei.
1987
. Метод объединения соседей: новый метод реконструкции филогенетических деревьев.Мол. Биол. Evol.
4
:406
–425Симмондс П., Л. К. Чжан, Ф. Макомиш, П. Балф, К. А. Лудлам и А. Дж. Браун.
1991
. Прерывистое изменение последовательности env-последовательностей вируса иммунодефицита человека (ВИЧ) типа 1 в плазменных вирусных и лимфоцит-ассоциированных провирусных популяциях in vivo: значение для моделей патогенеза {ВИЧ}.J. Virol.
65
:6266
–76Tajima, F.
1983
. Эволюционная взаимосвязь последовательностей ДНК в конечных популяциях. Генетика 105 : 437–460Вей, X., С. К. Гош, М. Э. Тейлор и др. (12 соавторов).
1995
. Вирусная динамика при инфекции вирусом иммунодефицита человека 1 типа. Nature 373 : 117–122Вольфс, Т. Ф., Г. Цварт, М. Баккер, М. Валк, К. Л. Куикен и Дж. Гоудсмит.
1991
.Встречающиеся в природе мутации в геномной РНК V3 ВИЧ-1 приводят к антигенным изменениям, зависящим от одной аминокислотной замены. Вирусология 185 : 195–205Ямагути Ю. и Т. Годжобори.
1997
. Эволюционные механизмы и популяционная динамика третьей вариабельной области оболочки ВИЧ в пределах отдельных хозяев. Proc. Natl. Акад. Sci. USA 94 : 1264–1269Zhang, L., R. S. Diaz, D. D. Ho, J. W. Mosley, M. P. Busch и A. Mayer.
1997
.Хозяин-специфическая движущая сила в эволюции вируса иммунодефицита человека типа 1 in vivo.J. Virol.
71
:2555
–2561Время генерации, продолжительность жизни, продолжительность возраста и частота линьки и их взаимосвязь с температурой у восьми видов кладоцер из реки Темза, показание
Андерсон, Б.Г .: Число предпозрелых возрастов, рост, относительный рост , и вариация в Daphnia magna .Биол. Бык. мар. биол. Lab., Woods Hole 63 , 81–98 (1932)
Google ученый
Андерсон, Б.Г., Дженкинс, Дж. К.: временное исследование событий продолжительности жизни Daphnia magna . Биол. Бык. мар. биол. Lab., Woods Hole 83 , 260–272 (1942)
Google ученый
Андерсон Б.Г., Люмер Х., Зупанчич Л.Дж .: Рост и изменчивость Daphnia pulex .Биол. Бык. мар. биол. Lab., Woods Hole 73 , 444–463 (1937)
Google ученый
Банта, А. М .: Исследования физиологии, генетики и эволюции некоторых Cladocera. Carnegie Inst. Wash. Publ. № 513 (1939)
Берри, А.Д .: Продуктивность реки Темзы в Ридинге. Symp. зоол. Soc. Лондон. 29 , 69–86 (1972a)
Google ученый
Берри, А.D .: Наличие и состав сестона в Темзе и роль детрита как источника энергии для вторичного производства в реке. Mem. Ist. Ital. Идробиол. (Дополнение) 29 , 473–483 (1972b)
Google ученый
Боттрелл, Х. Х .: Взаимосвязь между температурой и продолжительностью развития яиц у некоторых эпифитных Cladocera и Copepoda из реки Темзы, Рединг, с обсуждением температурных функций.Oecologia (Berl.) 18 , 63–84 (1975)
Google ученый
Боттрелл, Х. Х .: Соотношение между длиной и массой и ростом Cladocera из реки Темзы, Рединг. В процессе подготовки
Коркетт, К. Дж., Макларен, И. А .: Взаимосвязь между скоростью развития яиц и более старыми стадиями веслоногих ракообразных. J. mar. биол. Жопа. Великобритания 50 , 161–168 (1970)
Google ученый
Феррари, И.: Accrescimento Individual e ciclo stagniole della Daphnia middendorffiana di un Lago Alpino. Mem. Ist. Ital. Идробиол. 26 , 41–83 (1970)
Google ученый
Фрей Д.Г .: Сравнительная морфология и биология трех видов Eurycercus (Chydoridae, Caldocera) с описанием Eurycercus macracanthus sp. ноя Int. Revue ges. Hydrobiol. 58 , 221–267, 593 (1973)
Google ученый
Грин, Дж.: Размер и размножение Daphnia magna (Crustacea: Cladocera). Proc. зоол. Soc. Лондон. 124 , 535–545 (1954)
Google ученый
Грин, Дж .: Рост, размер и размножение Дафний (Crustacea: Cladocera). Proc. зоол. Soc. Лондон. 126 , 173–204 (1956)
Google ученый
Grygierek, E .: Некоторые данные о роли пищи в биологии Eudiaptomus zachariasi Poppe.Ekol. pol. 19 , 277–293 (1971)
Google ученый
Холл Д. Дж .: экспериментальный подход к динамике естественной популяции Daphnia galeata mendotae . Экология 45 , 94–112 (1964)
Google ученый
Hillbricht-Ilkowska, A., Patalas, K .: Методы оценки продукции и биомассы и некоторые проблемы методов количественного расчета зоопланктона.Ekol. pol, сер. B 13 , 139–172 (1967) (перевод как NLL RTS 6532)
Google ученый
Hrbáčková, M .: Распределение размеров новорожденных и рост Daphnia hyalina Leydig (Crustacea, Cladocera) из озера Маджоре в лабораторных условиях. Mem. Ist. Ital. Идробиол. 27 , 357–367 (1971)
Google ученый
Hrbáčková-Eslová, M.: Постэмбриональное развитие кладоцер. I. Группа дафний pulex . Жилет. čsl. Spol. зоол. 26 , 212–223 (1962)
Google ученый
Ингл, Л., Вуд, Т. Р., Банта, А. М .: Исследование продолжительности жизни, роста, размножения и частоты сердечных сокращений у Daphnia longispina в зависимости от ограничений в количестве пищи. J. exp. Zool. 76 , 325–352 (1937)
Google ученый
Кинне, О.: Рост, частота линьки, сердцебиение, количество яиц и время инкубации в Gammarus zaddachi в различных средах. Ракообразные 2 , 26–36 (1961)
Google ученый
Коринек, В .: Эмбриональное и постэмбриональное развитие Daphnia hyalina Leydig (Crustacea, Cladocera) из озера Маджоре. Mem. Ist. Ital. Идробиол. 26 , 85–95 (1970)
Google ученый
MacArthur, J.W., Baillie, W. H. T .: Метаболическая активность и продолжительность жизни. I. Влияние температуры на продолжительность жизни Daphnia magna . J. exp. Zool. 53 , 221–242 (1929)
Google ученый
Манн, К. Х., Бриттон, Р. Х., Ковальчевски, А., Лэк, Т. Дж., Мэтьюз, К. П., Макдональд, И.: Производительность и поток энергии на всех трофических уровнях в Темзе, Англия. В: З. Каяк, А. Хиллбрихт-Ильковска, ред., Проблемы продуктивности пресных вод, с.579–596. Варшава-Краков: Польские научные издательства 1972 г.
Google ученый
Майкл Р.Г .: Сезонные явления в естественной популяции кладоцера Ceriodaphnia cornuta Sars и наблюдения за его жизненным циклом. J. zool. Soc. Индия 14 , 211–218 (1962)
Google ученый
Муруган Н., Сиварамакришнан К. Г .: Биология Simocephalus acutirostratus King (Cladocera: Daphniidae) — лабораторные исследования продолжительности жизни, возраста, яйценоскости, роста и стадий эмбрионального развития.Freshwat. Биол. 3 , 77–83 (1973)
Google ученый
Navaneethakrishnan, P., Michael, R.G .: Производство яиц и рост Daphnia carinata King. Proc. Индийский акад. Sci. 73 , 117–123 (1971)
Google ученый
Смирнов Н.Н .: Eurycercus lamellatus (Chydoridae: Cladocera): полевые наблюдения и питание.Hydrobiologia 20 , 280–294 (1962)
Google ученый
Смирнов Н.Н .: Pleuroxus (Chydoridae): полевые наблюдения и рост. Hydrobiologia 23 , 305–320 (1964)
Google ученый
Wegleńska, T .